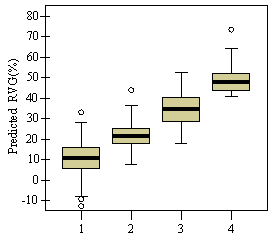|
 |
 |
 |



Working Paper 4
Factors Influencing Understory Douglas-fir Vigor in Multi-Cohort Prairie Colonization Stands at Fort Lewis, WashingtonReport to Fort Lewis Forestry Program
Derek Churchill
Rural Technology Initiative ACKNOWLEDGEMENTSI would like to acknowledge and thank the research partners involved in this study: Fort Lewis Forestry Program U.S.D.A. Forest Service Pacific Northwest Research Station College of Forest Resources, University of Washington Funding for this project was provided by U.S. Army Fort Lewis and U.S.D.A. Forest Service, Pacific Northwest Research Station through a Cooperative Agreement (PNW 04-CA-11261993-132) with the Rural Technology Initiative, College of Forest Resources, University of Washington.
ABSTRACTForest stands on Fort Lewis, Washington are being
managed for multiple values under an uneven-age silvicultural
system that
relies on natural regeneration. In stands that were former
prairies and have well drained, outwash soils, Douglas-fir
(Pseudotsuga menziesii) is the only understory conifer present
and the principal regeneration species. Managers wanted an
assessment of the vigor of the existing Douglas-fir regeneration
to determine whether it is a viable source for replacement
trees as well as management recommendations to ensure that
the silvicultural system is sustainable. Factors influencing
the vigor of Douglas-fir advanced regeneration were investigated
in thirteen stands at both the individual tree and stand
levels. Live crown ratio, height growth, height-to-diameter ratio,
and crown density were used to produce two methods of quantifying
vigor: a regression model that predicts volume growth as
a
percent of maximum site potential (relative volume growth)
and a simple vigor classification system. Overstory recruitment
potential and release potential were estimated for different
classes of vigor. At the individual tree level, understory
Douglas-fir with low levels of understory competition was
found to require an average 45% full sunlight or overstory stocking
of less than 150 SDI (30% full site occupancy (Long 1985))
to achieve vigor levels where recruitment into the overstory
without further release is likely. Between 10-35% full sunlight
or 150-275 SDI (30-55% full site occupancy), regeneration
was
found to be growing slowly but able to maintain its ability
to respond to release and regain growth rates comparable
with trees that were not suppressed, especially if less than
5m
in height at time of release. Below 10% full sunlight or
above 275 SDI (55% full site occupancy), regeneration was scarce
and of very poor vigor. Regeneration with high levels of
understory
competition was found to require more light to achieve the
same growth rates, and this effect increased in higher light
environments. A stand level model was developed and demonstrated
that while overstory density is the dominant factor influencing
understory vigor, understory stocking, shrub cover, spatial
arrangement of the cohorts are also important. A three stage
progression of overstory treatment types is recommended to
balance the tradeoffs between stand volume growth, structural
and habitat goals, and understory vigor. By combining elements
of shelterwood, group selection, and single tree selection
systems, multi-cohort, structurally complex stands can be
created and maintained in a shifting mosaic of patches. Results
suggest
that uneven-age management is possible with Douglas-fir on
dry sites, although stands will be structurally different
from west-side, late-successional forests that contain shade
tolerant
conifers and will require periodic stand entries to maintain. Table of ContentsList of Figures
List of Tables
SUMMARYIn west side forests in the Pacific Northwest, there is growing interest in uneven-aged management for ownerships with a mix of financial, wildlife habitat, and other objectives. Forest stands on Fort Lewis, Washington are being managed for such a mix of objectives under an uneven-age silvicultural system that relies on natural regeneration. Approximately 25% of the forestland (15,000 ac) at Ft. Lewis is dry-site Douglas-fir forest that established over the last 150 years on former prairies with coarse textured, droughty outwash soils. These stands have been managed for the past several decades with light thinning entries that remove 15-20% of standing volume at roughly 10 year intervals (Foster and Shaff 2003). In the understory, Douglas-fir is well established and is the only conifer species present. Ft. Lewis managers are relying on this naturally regenerated Douglas-fir to become overstory trees in the future as they continue the 10 year cycle of thinning entries. Managers wanted an assessment of the vigor of the existing Douglas-fir regeneration to determine whether it is a viable source of replacement trees as well as management recommendations to ensure that the silvicultural system is sustainable. Factors influencing the vigor of Douglas-fir regeneration were investigated at both the individual tree and stand levels. Sampling methods consisted of two phases. In the first phase, thirteen stands that represented the range of stand conditions in prairie colonization stands at Ft Lewis were inventoried using techniques that can be easily integrated into the existing inventory system. Variable radius plots and percent open sky estimates were used to measure overstory stocking and fixed area plots were used for understory trees and shrub cover. Measurements to determine live crown ratio (LC ratio), height-to-diameter ratio (HD ratio), height growth, and crown density rating were taken on understory trees. Competition from neighboring understory trees and shrubs was also measured by a percent crown overlap method that estimated the percent of projected crown area occupied by competitors. The second phase consisted of more detailed measurements and analysis on a smaller sub-sample of trees from the first phase to investigate the factors influencing understory vigor in greater depth, develop regression models that could be extrapolated to the larger phase I dataset, and to test and calibrate visual estimation techniques used in the phase I stand inventory with instrument based measurements. Two different methods were developed to assess the vigor of Douglas-fir regeneration. The first method consisted of the calculating the past 5 years of volume growth as proportion of open grown, maximum potential volume growth. Volume growth data from 19 open grown, “best” trees growing on the same soil type were used to determine maximum potential volume growth for a tree of a given height. This method, relative volume growth (RVG), allows for vigor assessment across a wide range of size classes and provides managers with a concrete measure of how understory trees are growing compared to the maximum potential for that site. A regression model was developed from the intensively analyzed trees in phase II to derive RVG from the growth metrics measured in the larger phase I stand inventory: height growth, HD ratio, LC ratio, and crown density rating.
Understory tree vigor was also assessed via a classification system that uses thresholds of height growth, HD ratio, and LC ratio to place trees into four vigor classes (Table S1). This simpler system was able to predict RVG with a degree of accuracy that is appropriate when general and rapid stand assessments are desired and a continuous vigor metric is not needed. The system can be easily integrated into most stand exam, marking, or cruising protocols. With some training, field technicians can quickly classify trees in the field using only visual estimates of these growth metrics, or by taking height and diameter measurements and visually estimating height growth and live crown ratio. The management implications of different levels of vigor were then defined by linking each of the four vigor classes with estimates of release potential and the likelihood of recruitment into the overstory without further release. Based on the average height growth and radial growth rates of trees in each vigor class (Table S1), it is very likely that most of the Douglas-fir regeneration found in this study will need release from overstory removal in order to recruit into the overstory (vigor classes 1-3). Although understory Douglas-fir appears able to maintain slow growth rates, reasonable LC ratios and HD ratios, and release potential in low light environments for several decades, it is unlikely that it can persist at slow growth rates in the understory until the overstory is thinned through natural mortality. Understory trees from vigor class one are currently at some risk of mortality from suppression and growth rates of trees from higher vigor classes will decline as the overstory closes over time. Table S1: Summary data for vigor classification system for understory trees. Trees must meet thresholds for all three growth metrics to be in a class, otherwise they are placed in the next lowest class.
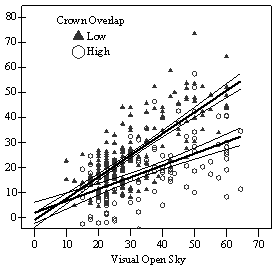
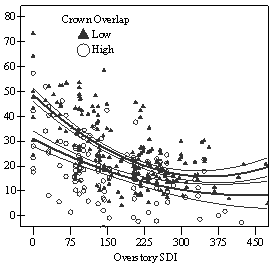
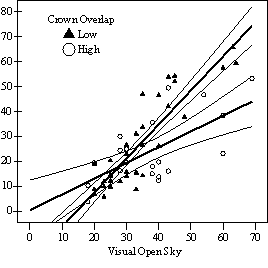
If understory trees are released to replace the overstory, evidence from this study and all other studies reviewed suggest that Douglas-fir will respond (Seidel 1983a, Helms and Standiford 1985, Oliver 1985, McCaughey and Ferguson 1988, Carlson and Schmidt 1989, Tesch and Korpela 1993, Deisenhofer 2000, Wright et al. 2000, Miller and Emmingham 2001, Kneeshaw et al. 2002). In all but the most severely suppressed trees, released trees appear able to regain growth rates comparable with trees that were never suppressed, recover from high HD ratios, and become viable replacement trees. The lag time before response, degree of response, and decline in HD ratios after release will depend on the level of overstory removal, the degree of understory competition after release, and pre-release live crown ratio and growth rates. Furthermore, risk of stem failure from high HD ratios and lag time before response to release appears to increase with increasing tree height. Trees taller than 5m in both vigor class one and two should be carefully evaluated to determine if they are worth releasing. A LC ratio of at least of 50% and an HD ratio of not more than 80 (measured at 15cm above ground) are suggested thresholds to guide this evaluation. Trees taller than 10m, especially from vigor class 1, are unlikely to be worth releasing. At the individual tree level, a significant relationship was observed between relative volume growth and overstory density, as measured by both percent open sky and Stand Density Index (SDI) (Figure S1). Regeneration with low levels of understory competition (less than 50% crown overlap) was found to require an average of 35-45% full sunlight or overstory stocking of less than 150 SDI (24 Curtis Relative Density (Curtis 1982); 30% full site occupancy (Long 1985)) to achieve higher classes of vigor where recruitment into the overstory without further release is possible (vigor class 3-4). If released, these higher vigor trees will quickly attain growth rates comparable with trees that were never suppressed, if they are not already there. Between 15-35% full sunlight or 150-275 SDI (24-44 Curtis Relative Density; 30-55% full site occupancy), regeneration was found to be growing too slowly to be able to recruit into the overstory without release (vigor class 1-2). However, understory trees did appear able to maintain the ability to respond to release and regain growth rates comparable with trees that were not suppressed, especially if less than 5m in height at time of release. Below 15% full sunlight or above 275 SDI (44 Curtis Relative Density; 55% full site occupancy) regeneration was scarce and of very poor vigor. Trees with high levels of understory competition (greater than
50% crown overlap) require more light to achieve the same vigor
levels, and this effect increases in higher light environments.
Only in very high light environments do they reach vigor class
3 and 4. Even in low light environments, negative effects on HD
ratios and live crown ratios are apparent. Without release from
crown crowding, trees with greater than 50% crown overlap will
suffer serious losses in potential volume growth. Pre-commercial
thinning is recommended for crop trees that have crown overlap
over 50% or are likely to before the next stand entry. Pre-commercial
thinning should be done after overstory thinning treatments as
harvesting will damage a proportion of crop trees.
To balance the tradeoffs between total stand volume growth, structural and habitat goals, and vigor of regeneration, patchy or spatially heterogeneous stands can be created and maintained in a shifting mosaic of patches. A three stage progression of overstory treatment types is recommended for patches within a stand. First, some patches can be managed to maximize volume growth in the overstory for one or more stand entries with light thinning from below treatments that keep stocking levels over 275 SDI. Although some regeneration may establish after these light thinning entries, it should not be a primary treatment objective as regeneration establishment will be a focus of the second stage. In the second stage, the overstory should be opened up enough (150-275 SDI) for regeneration to establish and maintain at an average vigor of class two. Depending on stand conditions and the objectives of the landowner, overstory trees can be removed across all diameter classes or from below, and small gaps can be created. These two cohort patches can be maintained for several stand entries by progressively reducing overstory density towards 150 SDI which will allow the regeneration to keep growing, albeit slowly, and maintain its release potential. Depending on how vigorous the regeneration cohort is, the patch should be moved to the third stage once the regeneration is between 5-10m in height. In the third stage, overstory SDI should be reduced below 150 to provide enough light for regeneration to grow vigorously and recruit into the overstory. Removing all the overstory trees in one entry will maximize growth of the regeneration cohort. However, if a multi-layered canopy is a management goal, the overstory can be removed over several stand entries and some overstory trees can be indefinitely retained for long term structure. Clumped retention, rather than dispersed retention, should be used as it provides more micro-sites with high light levels for the same amount of overstory SDI (Bailey and Tappeiner 1998, Lieffers et al. 1999, Coates et al. 2003, Drever and Lertzman 2003). A third cohort may establish after scarification from successive stand entries in areas of the patch that are not stocked with regeneration. Once the main regeneration cohort reaches commercial size, the patch can be moved back to the first stage and begin the cycle again. If some members of the original overstory cohort are retained, a three cohort structure is likely to be achieved in the next cycle if it is not already present.
By ensuring that different patches within a stand are at different stages in the progression, a horizontally and vertically complex stand with multiple cohorts will be created. Minimum patch size should be 0.25 ha (0.6 ac), as this appears to be the minimum gap size necessary for Douglas-fir to achieve high vigor levels in gaps (Ketchum 1994, Mailly and Kimmins 1997). Larger patch sizes, however, will be easier to manage. By varying patch size and the spatial arrangement of patch stages, a high degree of structural complexity can be created. Active management of the regeneration cohort should be also considered during the second and third stages of overstory treatments. As described above, pre-commercial thinning and shrub control around crop trees has the potential to significantly increase vigor levels. While natural regeneration appears to be providing sufficient stocking under thinned understories, gaps captured by shrubs and grasses may need shrub control and or planting to ensure timely stocking and full utilization of growing space. Gaps occupied with native shrubs provide important wildlife habitat, but many gaps are filled with non-native Scotch Broom (Cytisus scopariu) in the stands inventoried in this study. Prescriptions for individual stands will need to take into account the current vigor, height, crown crowding, spatial arrangement, and stocking levels of both overstory and understory cohorts, as well as shrub cover. Relatively uniform, even-age stands with high overstory SDI and little regeneration will need to be thinned to different densities throughout the stand to create a mix of stage one and two patches. Stage three patches can be added if large gaps are desired for wildlife habitat. Stands that already have a high level of patchiness can be treated by moving different patches to different stages depending on the stocking, vigor and height of the regeneration in each patch. Stands with overstory stocking below 150 SDI and high levels of understory competition will likely require a greater emphasis on pre-commercial thinning over further overstory removal to move the regeneration into vigor classes three and four. To provide an approximate assessment of stands at the patch level, inventory data can be broken down by plot to show the distribution of overstory SDI and understory trees per acre, average height, relative volume growth, and crown overlap. The results of this study support the conclusions of other investigators that uneven-age management is possible with Douglas-fir on dry sites using two-aged shelterwood systems (Seidel 1983b, Tesch and Korpela 1993), group selection (Coates and Burton 1997), or single tree selection systems that include heavy thinning at some point (Deisenhofer 2000, Miller and Emmingham 2001). By combining elements of all three systems, structurally complex, multi-cohort stands can be created that also produce significant wood volume. These stands will be structurally different from west-side, late-successional forests that contain shade tolerant conifers, however, and require ongoing, periodic thinning to maintain.
INTRODUCTIONIn west side forests in the Pacific Northwest, there is growing interest in uneven-aged management for ownerships with a mix of financial, wildlife habitat, and other objectives. Forest stands on Fort Lewis, Washington are being managed for such a mix of objectives under an uneven-age silvicultural system that relies on natural regeneration. Approximately 25% of the forestland at Ft. Lewis is dry-site Douglas-fir forest that established over the last 150 years on former prairies with coarse textured, droughty outwash soils. These stands have been managed for the past several decades with light thinning entries that remove 15-20% of standing volume at roughly 10 year intervals (Foster and Shaff 2003). In the understory, Douglas-fir is well established and is the only conifer species present. Ft. Lewis managers are relying on this naturally regenerated Douglas-fir to become overstory trees in the future as they continue the 10 year cycle of thinning entries. Considerable debate exists in the scientific literature and among foresters whether uneven-age management of Douglas-fir, which is moderately shade tolerant (Issac and Dimock 1958, Herman and Lavender 1990), is possible using single tree selection or whether group selection is necessary (Curtis 1998, Malcolm et al. 2001). Single tree selection typically involves removing individual mature and low-vigor trees dispersed throughout the stand in relatively frequent light thinnings with the goal of creating and maintaining multiple cohorts over time (Smith et al. 1997). While establishment of Douglas-fir seedlings in the understory is not uncommon after light thinning entries (Bailey and Tappeiner 1998, Harrington et al. 2003), growth rates generally decline substantially as the sapling stage is reached (1-3m, 3-10ft), virtually eliminating recruitment into the mid- and upper-canopy (Herman and Lavender 1990, Williams et al. 1999). To maintain adequate vigor and recruit into the mid- and upper- canopy, understory Douglas-fir saplings require a substantial amount of growing space (Oliver 1995). Thus for single tree selection to work, it is recommended that the overstory be heavily thinned early in a stand’s development and then kept open through successive light thinnings (Bailey 1996, Emmingham et al. 2000, Brandeis et al. 2001b, Tappeiner et al. 2002).
In group selection, trees are removed in patches of varying sizes in multiple thinning entries over time. This creates a mosaic of even-aged groups of differing ages within a stand (Smith et al. 1997). As group selection creates larger openings than single tree selection, it is often proposed as a more viable method for regenerating and growing shade-intolerant species (Coates and Burton 1997, Smith et al. 1997). Minimum gap size suggested for Douglas-fir is approximately 0.25-0.5ha (approximately 0.5 -1 ac) (Ketchum 1994, Mailly and Kimmins 1997) or a gap diameter of 1.5 times the height of the surrounding overstory trees (Malcolm et al. 2001). The critical question in making either approach work is the amount of growing space Douglas-fir needs to maintain adequate vigor in the understory, and thus at what densities and spatial arrangements to maintain the overstory. Numerous investigators have addressed this question by correlating measures of overstory density and/or light availability with understory Douglas-fir growth, both at the stand and individual tree level (Table 1). From these studies, a very general conclusion can be drawn that a maximum of 40% of full stocking or 30-40% of full sunlight is necessary to maintain adequate vigor. However, conclusions from different studies have been variable and can be difficult to develop into comprehensive silvicultural guidelines for several reasons. First, the definition of adequate vigor is often not well defined or consistent among studies. Many authors directly or indirectly define adequate vigor as growth rates that are close enough to open grown, maximum site potential for a tree to recruit into the overstory without any release in the future (Wampler 1994, Bailey 1996, Brandeis et al. 2001b, Drever and Lertzman 2001) Yet, in uneven-age management systems with periodic overstory removals, understory trees need only to maintain sufficient leaf area, stem stability, and root system to avoid mortality and be able to respond to release in the future (O'Hara 1996, Ruel et al. 2000). Thus growth rates that are far below open grown, maximum site potential can be considered adequate for understory trees. Clearly defining vigor in terms of a tree’s ability to grow into the overstory without further release, maintain its release potential, or merely survive is a critical step in translating the variable results created by the different definitions of adequate vigor (Table 1) into guidelines for uneven-age management.
Defining vigor in terms of maintaining release potential requires an understanding of response to release. Results from studies on release of Douglas-fir, as well as inference from release studies of other western conifers, suggest that Douglas-fir can respond to release after even severe suppression and that risk of mortality, lag time before response, and post-release growth are strongly related to duration of suppression, and pre-release growth rates and live crown ratio (LC ratio) (Helms and Standiford 1985, Carlson and Schmidt 1989, Tesch and Korpela 1993, Kobe and Coates 1997, Deisenhofer 2000, Wright et al. 2000). Thus, acceptable levels of risk and desired post-release growth rates should guide definitions of pre-release vigor in terms of thresholds for pre-release growth rates or live crown ratio (Helms and Standiford 1985, Tesch and Korpela 1993, Ruel et al. 2000). Several investigators have designed vigor classification systems that have set such thresholds for Douglas-fir (Carter and Klinka 1992, Emmingham et al. 2000, Miller and Emmingham 2001). However, these systems are based on different growth metrics, involve qualitative criteria that can be difficult to replicate, or were designed for different size classes of trees. As rates of height and radial growth are affected by tree size, vigor thresholds set for seedlings based on absolute, and not relative, growth metrics may not be appropriate for saplings. The same is true for height to diameter ratio (HD ratio). Although an HD ratio of 60 is a commonly used threshold of adequate vigor (Newton and Comeau 1990), this number is questionable as the stem height of diameter measurements and the total height of the tree influence the value and implication of the ratio (Mustard and Harper 1998, Wilson and Oliver 2000). More research is needed to develop thresholds for multiple growth metrics that can be applied across different size classes and correspond to definitions of vigor that have clearly defined management implications. The second reason for the lack of comprehensive guidelines for uneven-age management of Douglas-fir is the difficulty in accounting for the large number of factors that affect growing space in complex stands, often with methods that were designed for even-aged management systems. Traditional, distant-independent stem measurements such as basal area or Stand Density Index (SDI) (Reineke 1933) are often poorly correlated at the plot level to light levels reaching the understory (Chan et al. 1997, Brandeis et al. 2001a, Aukema and Carey 2003) and thus are typically weak predictors of understory growth for individual trees (Wampler 1994, Deisenhofer 2000, Brandeis et al. 2001a). Light or canopy closure measurements have shown greater predictive power (Carter and Klinka 1992, Chen and Klinka 1997, Drever and Lertzman 2001), but are time consuming and hard to translate into stand level management prescriptions. Accounting for side shading (Oliver and Larson 1996) from competition from neighboring understory trees and shrubs is also essential to better explain what controls the growth of advance regeneration (Brandeis et al. 2001b, Duchesneau et al. 2001, Canham et al. 2004). Adding further complexity is research that suggests that Douglas-fir may be more shade tolerant on drier sites (Carter and Klinka 1992, Wampler 1994, Bailey 1996, Chen et al. 1996, Williams et al. 1999). A final factor is tree size. There is evidence that as Douglas-fir get older and taller, its light requirements to maintain growth also increase (Carter and Klinka 1992, Messier et al. 1999). Growth models have been developed for multi-cohort stands in other forest types that factor in some or all of the complex set of variables listed above to help managers determine stocking guidelines for different stand structures and management objectives. These models include: stand-level, distant-independent models based on crown competition (Biging and Dobbertin 1995, Hasenauer and Kindermann 2002), leaf area (O'Hara and Valappil 1999), or SDI (Long 1995, Ralston et al. 2003); and spatially explicit crown and light models (Biging and Dobbertin 1995, Coates et al. 2003, Gersonde 2003). Many of these models show promise for uneven-age management of Douglas-fir. However, spatially explicit models in particular require substantial investments of resources to develop and more inventory information and technical resources than most management agencies typically have. There is a clear need for stand level growth models that do not require spatially explicit inventory information but can account for the vertical and horizontal heterogeneity of multi-cohort stands (Monserud and Robinson 2003). In this study, I inventoried vigor and stand structure of naturally regenerated Douglas-fir advanced regeneration in prairie colonization stands at Ft. Lewis, Washington. Methods were developed to quantitatively assess and classify vigor using metrics that can be integrated with existing inventory datasets. I sought to clearly define the implications of different levels of vigor by linking quantitative classifications of vigor with estimates of release potential and the likelihood of recruitment into the overstory without further release. These estimates were made from some data gathered in this study, but primarily from results from other investigators. I then used these vigor assessment methods to investigate and model the factors influencing vigor of understory regeneration at the individual tree and stand levels. Finally, results were combined into silvicultural recommendations for uneven-age management in dry site Douglas-fir forests.
METHODSField sampling Study sites were restricted to forests that have colonized former prairies and have somewhat excessively drained, glacial outwash soils of the Spanaway and Fitch series (Typic Melanoxerands) (Anderson et al. 1955, Foster and Shaff 2003). Thirteen stands were selected for the phase I stand inventory using a proportional, stratified random sample to produce a set of stands that were a good representation of the range of structural conditions found in the total population of prairie colonization stands at Ft. Lewis (Foster, pers comm.). All stands had at least two thinning entries. Site index (King 1966) ranged from 34-38 (metric) (110-125, English) (Foster, unpublished data) and was assumed to be 35 (metric) (115; English) for all stands given that within stand variation is as great as among stands due to the heterogeneity of the glacial soils. In each stand, a systematic grid with a random start was used to locate a pre-determined number of plot centers. The number of plots was based on the size and basal area class of the stand. In some large stands, only a portion of the stand was sampled. Three sets of measurements with different plot configurations were taken at each plot center: (1) shrub cover and dead understory trees, (2) overstory trees, and (3) live understory Douglas-fir trees. Percent cover of shrubs taller than 1.37m (4.5ft) and less than 1.37m was estimated using a 0.008ha (0.02ac) fixed area plot. The number of dead understory trees was also tallied in this plot. The overstory was sampled in two ways. First, trees over 15cm (6in.) diameter at breast height (dbh; diameter at 1.37m above ground level) were sampled using point sampling (variable radius plots) (Bitterlich 1947). Basal area factors used were 28, 34, or 40 (English scale) depending on the basal area class of the stand. Second, a visual estimate of the percent of open sky (VOS), excluding shrubs or trees under 15cm (6in.) dbh, was made from each plot center. The same person estimated VOS in all 13 plots. This VOS measure was modified from Brandeis et al. (2001a) and Deisenhofer (2000) and consisted of estimating the percent openness of the canopy in both the northern and southern halves of the sky and averaging them. Where VOS was at least 20% higher in the southern half, the average was weighted to adjust for edge effects (Hasenauer and Kindermann 2002) using the equation below. It is treated as a unitless, relative measure, instead of an absolute measure of percent of open sky.
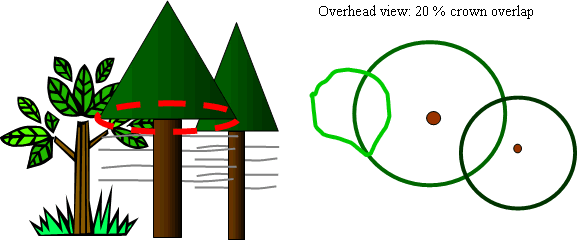
Eq. 1: Weighted VOS = (3VOS,South + VOS,North)/4 . Understory trees, defined as less than 15cm (6in) dbh and taller than 1.37m (4.5ft), were sampled within a circular fixed area plot of either 0.04 or 0.008ha (0.1 or 0.02ac), depending on the BA class of the stand. For all understory trees, dbh was measured and crown class determined based on the height of trees within a cohort: dominant, co-dominant, intermediate, or overtopped. Crown class for open grown trees not near clumps was determined by comparing the height of the tree to nearby trees in the same cohort. The following additional measurements were taken for all understory trees, or a sub-sample, depending on the number trees in the plot: diameter at 15cm (6in) above ground; total height; height to live crown, measured at the lowest whorl with two live branches with new growth; annual height increment for each of the past six years; total age determined from whorls and bud scars, crown density, and degree of crown crowding from intra-cohort competition (neighboring understory trees) and shrubs. Crown density was measured with a 1-5 rating that combined visual estimates of average branch length and diameter, number of internodal buds and branches, and number of branches and each whorl; all in the last 3 years of growth. Ranges of these variables used to rate trees are provided in Appendix A. Understory competition or crown crowding, defined as any infringement on the live crown of the sample tree from neighboring understory trees in the same cohort or shrubs, was measured with a percent crown overlap method. At the base of the live crown, a horizontal, two dimension circle was visualized around the tree using the longest live branch as the radius. The proportion of this two dimensional circle (projected crown area) overlapped by foliage of neighboring trees or shrubs from the base of the live crown to the top was estimated to the nearest 10 percent (Figure 1).
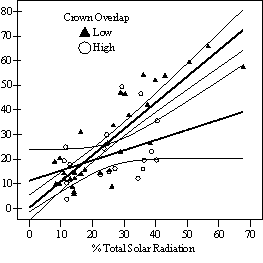
Data from the phase I stand inventory were generated from 13 stands, 212 plots, and 637 understory Douglas-fir trees and used to broadly characterize structure of the overstory, understory, and shrub layers. After this analysis, a sub-sample of 25 plots from the 212 phase I plots was selected for further intensive analysis. A stratified random sample was used to ensure that the phase II sub-sample of 25 plots came from a balanced distribution of light environments. At each phase II plot, two understory trees were selected for destructive sampling: the tallest tree within the cohort and the tree closest to the average height. For each tree, dbh of all overstory trees were recorded in a variable radius plot centered on the sample tree. Before each tree was cut down, the percent crown overlap method from phase I was expanded to include a vertical dimension. For each competitor, the two dimensional horizontal overlap and the percent overlap along the vertical live crown of the sample tree were estimated. By adding all the competitors together, an estimate was made of the percent occupancy of competing understory trees and shrubs within a three dimensional cylinder projected from the base of the live crown to the top of the tree with the radius being the longest live branch. After each tree was cut down, percent open sky was estimated using the VOS procedure described above and a hemispherical photograph was taken as close to 66% of tree height as could be safely reached with a 5m (16.4ft) orchard ladder using a digital Nikon camera with a 2.5 F-Stop fisheye lens mounted on a monopod with a level. Gap Light Analyzer/C, version 2.0 light modeling software (Frazer et al. 1999) was used to analyze these photographs and determine site openness and an index of total solar radiation (TSR) (Canham et al. 1990, Frazer et al. 1997). TSR combines the seasonal distribution of the sun’s path with the distribution of canopy openness to calculate a single index of available light in units of percentage of full sun for a specified growing season which was set at April 1 – Oct 15th for the Ft Lewis area. Height growth increments were measured from the top of the tree to breast height and checked against rings counted on a disc cut at dbh. Discs were also cut at ground level and at the base of the live crown. These discs were later oven dried for 48 hours, sanded, and analyzed using a high resolution computer scanner and WinDENDRO v2001a (Regent Instruments, Inc.) tree ring analysis software, which calculated age and annual radial growth along 4 radii for each disc. These radial growth measurements were combined with height growth data to calculate annual volume increment using a tapered rocket formula where the three discs were used to create two tapered cylinders and a cone at the top. Inside bark diameters were used. Height to diameter ratio history was also calculated using the ground level disc, but diameters were adjusted for bark thickness, drying, and the discrepancy between field measurements with a dbh tape and radial measurements on a disc. This was done by multiplying all inside bark diameters by the ratio of the 2004 field measured diameter to the 2004 inside bark diameter obtained from WinDENDRO.
Assessing vigor
For the 54 trees that were intensively analyzed in phase II, actual RVG was calculated for every year from when the tree was 1.37m to its current height. The last 5 years were averaged to create the primary vigor metric for further analysis: actual 5yr RVG. For the 637 understory trees in the stand inventory, however, coring trees to measure diameter and volume growth was intentionally not done to keep the sampling design within what could be practically integrated into the existing Fort Lewis inventory system. Exploratory analysis was thus done with the 54 tree data set to test which variables that were measured in the stand inventory best correlated with actual RVG. These variables were: past 5yr mean annual height growth increment (HG), 2004 live crown ratio (LC Ratio), 2004 height to diameter ratio (HD Ratio), and visual crown density rating (CDR). Single variables, as well as biologically meaningful products of these variables, were tested using simple linear regression. Stepwise, linear regression was then used to test multiple variables and derive a best fit model using SPSS, version 12.0 (SPSS 2003). This model was used to predict 5yr RVG for the 637 understory trees from the stand inventory. The second method of assessing vigor was a simple four class classification system. This additional method was developed to provide a more efficient and sufficiently accurate means for managers to assess vigor of understory cohorts, conceptualize vigor, and design silvicultural prescriptions. Classes were based on thresholds of HD ratio, HG, and LC Ratio that are listed in Table 2. Trees must meet the thresholds for all three metrics to be placed in a certain class or else they are placed in the next lowest class. Crown density rating was not included to allow for this vigor assessment method to be easily used with existing data sets. Also, crown density rating is a qualitative measure that requires consistency, which can be hard to achieve with multiple observers. Table 2: Summary data for vigor classification system for understory trees. Trees must meet thresholds for all three growth metrics to be in a class, otherwise they are placed in the next lowest class.
A HD ratio of 70, measured at 15cm above ground, was chosen as the HD threshold for high vigor (class 4) (Table 2). This correlates with an HD ratio of 60 at the root collar. This threshold value is based on observations from: Newton and Comeau (1990) who suggest that HD ratios over 60, measured at the root collar, threaten the long term growth potential of young plantation trees; Wonn and O'Hara (2001) who showed that an HD ratio of 80, measured at dbh, is a critical threshold for stem stability in interior Douglas-fir; Cole and Newton (1987) who found that decreases in height growth occurred at HD ratios above 70 in plantation trees; and Emmingham et al. (2000) who specified 60 as a threshold for a vigorous understory tree, 80 for a stable tree, and 100 for a weak tree. Height growth thresholds were based on approximately 66%, 33%, and 15% of the site index potential height growth for the average tree height found in the study (King 1966). Live crown ratio thresholds (Table 2) were based on the positive relationships of live crown ratio to post-release growth observed by Helms and Standiford (1985), Seidel (1983a), Oliver (1985), and Tesch and Korpela (1993); as well as recommendations by Emmingham et al. (2000). The two methods of assessing vigor were compared to evaluate whether
the simpler vigor classification system could estimate 5yr RVG
within a reasonable range. Analysis of variance (ANOVA) and a Scheffe’s
post hoc test was used to test for significant differences between
the mean RVG values for each vigor category, for both the phase
1 and phase 2 datasets ( Overstory recruitment and release potential Similar to Tucker and Foster (unpublished), the number of years needed to reach the average overstory height of the 13 stands, 45m (148ft), was used as an indicator of the likelihood of reaching the overstory without further release, assuming that current height growth rates are maintained and that trees are currently 5m (16.4ft) in height. This indicator was a simple thought exercise used to compared vigor classes and purposely ignored the dynamic nature of forest stands and the fact that Douglas-fir height growth begins to slow as trees reach approximately 25m (80ft) on site class III sites (King 1966).
As a biological limit exists regarding how slow trees can grow and remain alive (Oliver and Larson 1996, Kobe and Coates 1997), risk of mortality from suppression was then examined. As the 54 tree sub-sample was a representative sample of the total population of understory trees, the slowest radial growth rates found from analysis of lifetime radial growth rates were assumed to be close to the threshold of mortality. These minimum growth rates were compared with mortality threshold rates from other studies. Estimates of mortality risk for each vigor class were based on how close the past 5yr mean radial growth increment were to these mortality thresholds. HD ratio thresholds for each vigor class were also considered as high HD ratios are associated with a higher risk of stem failure and suppression mortality (Wilson and Oliver 2000, Wonn and O'Hara 2001). Release potential was defined in terms of a released tree’s ability to regain comparable growth rates, for a given light environment, to trees that did not undergo suppression. To assess release potential, lifetime RVG histories of the 54 intensively analyzed trees were examined for patterns of suppression and release. Trees that showed patterns of release were compared to trees growing in similar light environments that had not undergone suppression. Lifetime HD ratios was also examined to see the extent to which trees recovered from high ratios. Although the exact history of overstory removal around each tree could not be determined with a high degree of confidence, it was assumed that at least some of the trees currently growing in open conditions had been released in the past and should therefore show a growth response. Finally, the live crown ratio thresholds for each vigor class were also considered. Factors influencing vigor: individual tree and stand level At the stand level, two methods were used to compare the overall vigor of the understory between stands. First, the number of understory trees per acre in each vigor class was tabulated for each stand. Second, the stand level, mean tree 5yr RVG was calculated for all trees, and then separately for crop trees, by averaging the mean RVG of each plot with understory trees. A non-linear model was developed to explain variations in stand mean RVG and test which factors influence understory vigor at the stand level. The model was based on the concept that average growth of an understory cohort is determined by growing space available to the cohort divided by the number of trees in the cohort (O'Hara 1996, Oliver and Larson 1996). Although multiple understory cohorts did exist in some stands, all understory trees were lumped into one cohort to simplify the analysis. Stand density index (SDI) was chosen to measure growing space occupancy for both the overstory and understory, as it has been shown to be a good indicator of leaf area index (Long 1995, O'Hara 1996) and is commonly used to measure site occupancy (Long 1985, Long and Daniel 1990). Percent shrub cover was tested in the model to account for its potential effect on resource availability. The average crown overlap for the stand was also tested to account for the spatial distribution of understory trees; whether they are clumped or more evenly distributed. A final variable was tested to account for the distribution of understory trees in relation to the heterogeneity of overstory density. This variable was the ratio of average overstory SDI for plots with understory trees to the average overstory SDI of all the plots. Stands with understory trees in more open plots thus have a lower value than stands where more trees are in plots with higher levels of overstory or where gaps are occupied with shrubs instead of trees. Several model forms were tested and the best combination of variables was chosen through iterative trials. SPSS, version 12.0 (SPSS 2003) was used for this analysis.
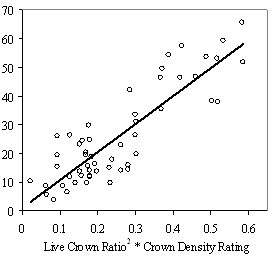
ResultsThe 13 stands inventoried had a wide range of stand structures and understory vigor levels (Table 3). Some were very open with large, evenly distributed overstory trees and dense understories and resembled shelterwood cuts. Others had large gaps and dense patches and were the result of ongoing prairie colonization and possibly some group selection harvesting. Yet others were fairly dense with small gaps. Overstory stocking ranged from very open (91 SDI, 20% of full site occupancy) to somewhat dense (298 SDI, 60% of full site occupancy), with 510 SDI being full site occupancy (Long 1985). Understory densities had an even greater range: 10-842 trees per acre. Only the most open stand had an average crop tree RVG above 35% and the majority of crop trees in vigor classes 3 and 4. The three stands with overstory SDI levels close to 150 had average crop tree RVG values under 30%. Assessing vigor Stepwise linear regression produced the following best fit model to predict 5 yr mean RVG using HD ratio, height growth increment, LC ratio, and crown density rating (CDR) and total height (Ht). The model produces negative RVG values for trees with very high height to diameter ratios and/or very low live crown ratios. Although negative values do not make biological sense, they were included in subsequent analysis as indicative of RVG close to zero. A second, slightly weaker model to predict RVG that does not include crown density rating is listed in Appendix B for use with data sets where crown density rating is not available. Eq 2: Predicted RVG = 1.337 + 0.076LCR2CDR - 0.009Ht(m) - 0.028ln(HD Ratio) + .003HG(cm)
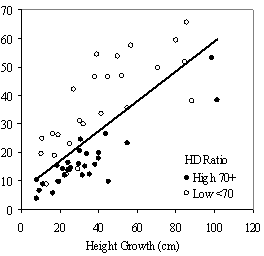
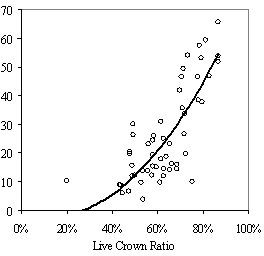
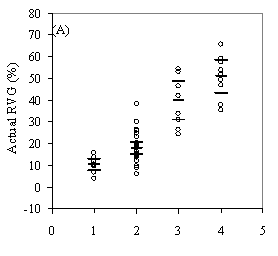
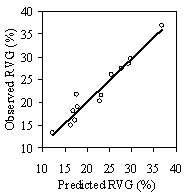
Table 3: Summary data for all 13 stands. Standard errors are in ( ) for selected variables. Values are stand level means unless otherwise stated. The second method of assessing vigor, vigor classes based on thresholds, is compared with RVG in Figure 4 for both phase I and II datasets. The extreme upper values in each category are due to high height to diameter ratios. The means of actual and predicted RVG for the 4 vigor categories are all significantly different from each other (p<0.05). The combined phase I and II average RVG values (rounded to the nearest 5%) for each vigor class are presented in Table 2.
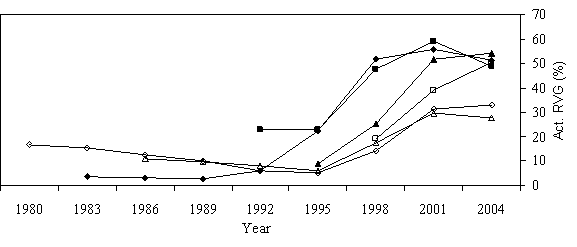
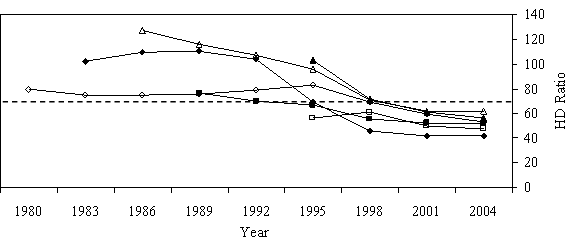
Overstory recruitment and release potential Of the 54 phase II trees, six were found to have strongly responded to release (Figure 5a). Pre-release RVG rates were under 20% and radial growth rates were less than 2mm (0.08in.) per year. Trees maintained these slow growth rates for up to 15 years and were all less than 5m (16.4 ft) in height at time of release. Their height to diameter ratios all declined to values below 65 and in some cases were over 100 prior to release (Figure 5b). These six trees had RVG rates similar to trees in the same light environment that had not undergone suppression and were all above the regression lines in models relating RVG to open sky (Figure 6a).
Factors influencing vigor: individual tree level
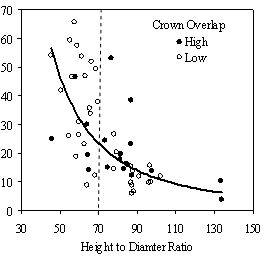
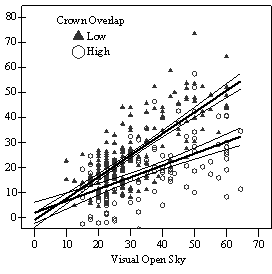
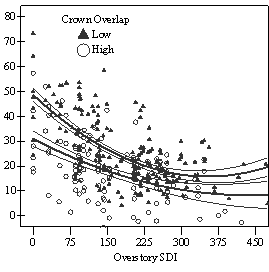
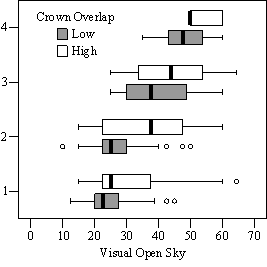
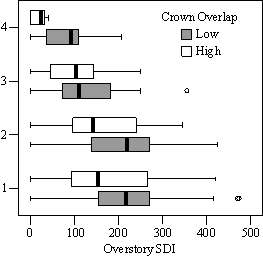
In the phase I VOS (Figure 7a), phase II VOS (Figure 6a), and phase II TSR (Figure 6b) regression models, ANCOVA showed statistically significantly steeper slopes and higher R2 values for trees with low crown overlap than trees with high crown overlap. Intercepts were not significant for either high or low crown overlap. The effect of crown overlap was not significant at the lowest light levels but became significant at the point where the 95% CI of the regression lines diverged, and had an increasing effect as open sky increased and overstory density decreased. Phase I and II regression models for SDI had similar slopes, intercepts and R2 values and so only the results for the phase I, larger data set are presented in Figure 7b. Low crown overlap had a higher R2 value and a significantly different intercept than high crown overlap, but the slope and curvature were not significantly different. Similar to the VOS models, a pattern of an increasing effect of crown overlap as SDI decreased was found. Between 200 and 250 SDI (Figure 7b), there is a clump of 6 trees above 35% RVG that were all on the edges of gaps and so were receiving more light for their SDI value than trees that were underneath a more continuous canopy. Accounting for this clump of outliers, there is a clear jump in the response of RVG at 150 SDI. It should be noted that 0 SDI does not mean that the tree is open grown, only that no overstory trees are within the plot radius measured. The second vigor assessment method, the 4 vigor classes, was also related to VOS, SDI, and crown overlap using boxplots (Figure 8). These were derived to clearly show the ranges of VOS and SDI values that trees from the 4 vigor classes were found in.
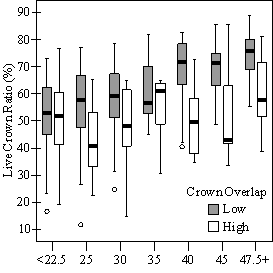
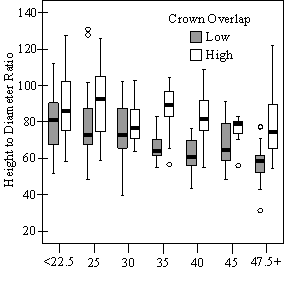
In addition to the main vigor metric, RVG, regression models predicting live crown ratio and HD ratio were developed from the same set of explanatory variables as RVG. LC ratio and HD ratio showed the same basic relationships to VOS, SDI and crown overlap, but models were consistently weaker than models for RVG and so are not presented here. Instead, boxplots are shown in Figure 9 to display the distribution of LCR and HD ratio data by VOS and SDI class and crown overlap.
Factors influencing vigor: stand level
Eq. 3:
Eq. 4:
Between 85-90% of the variation in RVG was explained with only overstory SDI, understory SDI, and tall shrubs or low shrubs in each model. Non-significant coefficients were included to add predictive power. As only 13 data points were used, the asymptotic confidence intervals of the non-linear regression method are large. With more data points, they may in fact be skewed in a negative or positive direction and be significant.
DISCUSSIONAssessing vigor Combining height growth, HD ratio, LC ratio, and crown density
rating into a regression model based on relative volume growth
proved to be a more powerful approach to both predicting current
volume growth and measuring response to overstory stocking and
crown crowding than using individual, absolute metrics. All of
the coefficients have signs that make biological sense. The significance
and negative sign of total height in the model reflects the change
in maximum potential height and radial growth and exponential increase
in volume growth with increasing tree size (King 1966). Other investigators
examining growth of understory Douglas-fir in relation to environmental
variables have generally used two or more vigor metrics in their
analysis (Carter and Klinka 1992, Mailly and Kimmins 1997, Williams
et al. 1999, Deisenhofer 2000, Drever and Lertzman 2001). However,
combining multiple crown and growth metrics into a single, relative
metric that is strongly correlated with volume growth is advantageous
in situations where coring large numbers of trees is not feasible
and investigators want a single, continuous, and comprehensive
vigor metric to assess and compare advanced regeneration across
a wide range of tree sizes.
Overstory recruitment and release potential There is a biological limit to how slow trees can grow and survive. Models from Kobe and Coates (1997) for western conifers in interior boreal forests show an exponential increase in probability of mortality as annual radial growth declines below a species specific, threshold level for a period of 4 years. If coastal Douglas-fir is assumed to have a shade tolerance in between lodgepole pine and hybrid spruce (Coates, pers comm.), these models suggest that probability of mortality in the next 3 years is approximately 30% at an annual radial growth rate of 0.5 mm per year. Although this extrapolation is clearly problematic, it approximates the slowest growth rates observed in this study (0.3mm) and by Deisenhofer (2000), whose slowest rates were 0.2-0.5 mm. While further research is needed to better quantify the risk of mortality associated with slow growth rates for coastal Douglas-fir and adjust for tree size, a rate of 0.5mm per year sustained for at least four consecutive years was considered the most reasonable estimate of a threshold of mortality risk. Vigor class one trees are at or close to this threshold and class two trees are not far away, while class three and four trees appear to have a low risk of mortality from suppression (Table 2). This is further evidence that class one and two trees will not recruit into the overstory without release.
Results from this study (Figure 5), and
all other studies reviewed on the release of Douglas-fir advanced
regeneration and other
western conifers, indicate that response to release, from even
severe suppression, is highly likely and risk of mortality
from thinning shock is low (Seidel 1983a, Helms and Standiford
1985,
Oliver 1985, McCaughey and Ferguson 1988, Carlson and Schmidt
1989, Tesch and Korpela 1993, Deisenhofer 2000, Wright et al.
2000, Miller and Emmingham 2001, Kneeshaw et al. 2002). Furthermore,
evidence from this study supports the conclusion reached by
Tesch and Korpela (1993) and Wright et al (2000) that, for
a given
light environment, released trees are capable of attaining
growth rates comparable to trees that did not undergo suppression.
Wright
et al (2000) also found that the time it takes to attain comparable
growth rates tends to increase with length and severity of
suppression, some trees may never attain such rates in cases
of severe suppression,
and trees can undergo multiple episodes of suppression and
release. After release, height growth often declines for a short period while trees build up the root capacity and stem conductive tissue needed for increased growth (Helms and Standiford 1985, Kneeshaw et al. 2002). The result can be a rapid decline in height to diameter ratios as was observed in this study and in other data collected from 103 understory Douglas-fir saplings at Fort Lewis (Foster, unpublished data). Foster (unpublished data) observed that mean HD ratios declined from 92 to 72 in 3 years after partial release. Emmingham et al. (2000) used an HD ratio of 60 at ground level, which corresponds to 70 at 15cm above ground, as a threshold for acceptable vigor of understory trees. This threshold, which is a common threshold for young plantations (Newton and Comeau 1990), does appear to be a reasonable guide for assessing overstory recruitment potential without release for understory Douglas-fir in this forest type. However, trees with higher HD ratios can recover if released and become viable replacement overstory trees. Furthermore, risks of stem failure associated with high HD ratios (Wonn and O'Hara 2001) are likely to be lower in understory environments where the overstory ameloriates the effects of heavy snow loads and high winds.
While it is highly probable that most of the understory trees surveyed in this study are capable of release, the lag time before response to release, degree of response, and decline in HD ratio after release will depend on the level of overstory removal, the degree of crown crowding after release, and pre-release growth rates, live crown ratio, and length of suppression. In a study of response to release after shelterwood removals in Douglas-fir stands, Tesch and Korpela (1993) showed that pre-release volume growth was strongly correlated with post release volume growth. In addition, height growth and live crown ratio have also been shown to be strong predictors of post-release growth for Douglas-fir and other western conifers. In a summary of vigor metrics that predict release potential, Ruel et al. (2000) recommends HD ratio and crown metrics such as number of buds and branches for intermediate shade tolerant species such as Douglas-fir. As RVG combines all of these metrics, it is reasonable to assume that RVG is a strong predictor of response to release. For a given post-release light environment, trees with higher pre-release RVG are likely to achieve higher post-release RVG and thus have a greater probability of attaining rates comparable to trees that did not undergo release and do so in a shorter period of time. Results from this study are not sufficient to statistically verify this conclusion, however, and further investigation is needed to confirm the value of RVG as a predictor of magnitude of, and lag time before, response to release. Tree height should also be factored into release potential. Trees observed in studies of release have generally been less 5m in height at time of release. Risk of stem failure from high HD ratios is greater in taller trees due to greater physical stress loads on the stem (Wilson and Oliver 2000, Wonn and O'Hara 2001). Rebuilding live crown and rebalancing HD ratios after release takes longer in taller trees as new growth is a smaller proportion of total tree size. While no height cutoff exists at which response to release is possible (Deisenhofer 2000, Wright et al. 2000), the window of risk of stem failure from high HD ratios and the lag time before response to release is likely to be longer for taller trees. Kneeshaw et al (2002) and Deisenhofer (2000) found some evidence of an negative effect of increasing tree height on the degree of response to release. In open grown plantations trees, Wilson and Oliver (2000) found that 10m was a critical height for trees to be able to recover from high HD ratios and reach full growth potential. Based on these findings regarding overstory recruitment and release potential, the following assessments were made regarding each vigor class. Class four trees are “free to grow” in that they have enough growing space projected into the future to have a high likelihood of reaching the overstory without further release. If they are released, either from overstory removal or pre-commercial thinning of neighboring competitors, they will respond quickly and attain growth rates close to or at the maximum potential for that light environment, if they are not already there. Class three trees can likely maintain their current growth rates for some time, but will probably need release from overstory removal or pre-commercial thinning of neighboring competitors at some point to recruit into the overstory. If released, they will quickly attain growth rates close to or at the maximum potential for that light environment. It is highly unlikely that class two trees will reach the overstory without release. They will need release from overstory removal, and possibly also from pre-commercial thinning of neighboring competitors, in the next 1-2 thinning cycles to avoid severe suppression and substantial risk of mortality. If thinned within this time period, they should attain post-release growth rates comparable to trees that were not suppressed, after a period of time. Class one trees are currently at some risk of mortality from suppression and will not reach the overstory without release. Although it is likely they will respond to release, they may take a long time to respond and some may never attain post-release growth rates comparable to trees that were not suppressed. Trees taller than 5m in both class one and two should be carefully evaluated to determine if they are worth releasing. A LC ratio of at least of 50% and an HD ratio of not more than 80 (measured at 15cm above ground) are suggested thresholds to guide this evaluation. Trees taller than 10m, especially from vigor class 1, are unlikely to be worth releasing. The ability of taller trees to attain post-release growth rates comparable to trees that were not suppressed is likely to be lower, and lag time before response may be so long that it may be faster to start with new regeneration. The assessments and height thresholds regarding overstory recruitment and release potential for the four vigor classes are conservative estimates based on incomplete information. Further research should be undertaken to empirically test these estimates to avoid unforeseen consequences.
Factors influencing vigor: individual tree level The fact that using a 30% 3-dimensional or 50% 2-dimensional threshold of crown overlap as an indicator variable performed better than using it as a continuous variable suggests that either understory trees can withstand a certain level of crown crowding before growth reductions occur, or that effects of small changes in competition levels are not detectable within the range of other factors influencing growth or with the measurement techniques used. Caution is advised in using the 2-dimensional crown overlap method. Occasionally a tree had lot of competition at the base of the live crown from short neighbors, but very little in the upper 2/3rds of the live crown. These trees, although relatively uncommon in this data set, had a high percent crown overlap but little crown crowding in the majority of the crown. A way to account for this and make the 2-dimensional method more precise would be to estimate percent overlap at the base of the live crown and also halfway up the live crown and then average the two estimates together. Also, one of the main sources of measurement error in any of these approaches is including dead neighbors that have no current effect but clearly influenced crown development of the sample tree in the past. Dead neighbors were not common in this study and were mostly found on the ground; and were not included in any analysis. Similar to other studies (Table 1), 45% full sunlight was found to be the average level where understory trees with little intra-cohort competition could achieve vigor levels where recruitment into the overstory without further release is likely (vigor class four) (Figure 6, 7, & 8). For vigor class three trees, this average is 35% full sunlight. For vigor classes one and two, the average was found to be 10-15% full sunlight. Below 10% full sunlight, regeneration was scarce and of poor vigor. The linear relationship of measures of light and RVG, and the wide scatter in the models, is consistent with other studies on Douglas-fir saplings for the range of light levels sampled (Williams et al. 1999, Deisenhofer 2000, Drever and Lertzman 2001). Although Drever (2001) used a non-linear regression model, the relationship of height growth and log radial growth to percent full sun is almost linear for the site index that is closest to the stands sampled in this study. He attributes the wide scatter around the best fit line and high variability in growth at a given light level to genetics, mycorrhizal associations, previous periods of suppression, disturbance history, competition for resources, morphological differences, and micro-site factors. The variance introduced by using the regression model to predict RVG, as well as measurement error, are undoubtedly other factors in this study.
The linear relationship of light to RVG should not be extrapolated to trees smaller than the 1.4m cutoff used in this study. Other authors investigating Douglas-fir seedlings have found that growth increases linearly to a certain light threshold and then declines in slope thereafter (Carter and Klinka 1992, Mailly and Kimmins 1997, St.Clair and Sniezko 1999). This pattern is common for more shade tolerant conifer species of all size classes (Wright 1998), and suggests that Douglas-fir may grow equally well under partial overstories as in full sunlight at the seedling stage, at least on harsh sites. Once seedlings are established and able to survive the summer drought, the asymptotic relationship then disappears. In mixed conifer forests of eastern Oregon and Washington, Seidel (1983) noted that Douglas-fir regeneration in clearcuts often failed and that the shelterwood system resulted in satisfactory regeneration. Plantations have been successfully established on prairie soils at Ft. Lewis (Foster, pers comm.). Natural regeneration, however, may have greater or equal survival and growth during the first few years after germination under partial canopies. The inferiority of stand level density measures over light measures at predicting growth responses of individual trees is consistent with findings that density measures are weaker predictors of growth (Wampler 1994, Deisenhofer 2000, Brandeis et al. 2001b, Page et al. 2001) and not well correlated with light (Chan et al. 1997, Brandeis et al. 2001a, Aukema and Carey 2003). Similar to this study, Wampler (1994) found a quadratic relationship between overstory basal area and percent of maximum height growth with a similar wide but clear band in the data and sharp increase at a 100 ft2/acre. This corresponds to roughly 125 SDI, which is slightly lower than the sharp increase in growth found at 150 SDI in this study (Figure 7b). These results indicate that 125-150 SDI (25-30% of full site occupancy, (Long 1985); 20-24 Relative Density (Curtis 1982)) represents a threshold where overstory density is low enough to begin to let enough light through for RVG to exceed 30% and create conditions for trees with a vigor category of 3 or 4. An upper threshold for class one and two trees appears to be around 275-300 SDI (54-59% full site occupancy; 44-48 Relative Density (Curtis 1982)) as less than 25% of class one and two trees were found above 275 SDI (Figure 8). These SDI thresholds are in line with conclusions from other studies (Table 1). Factors influencing vigor: stand level
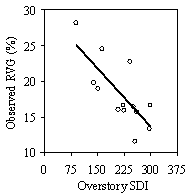
Overstory SDI is the dominant driver in the model confirming the basic idea that it is the main occupier of growing space (Oliver and Larson 1996). Understory SDI is also a significant factor. Stands with low overstory SDI but high understory stocking have lower than expected RVG. An extreme example of this is Recondo Johnson (Rj, Table 3). The portion of this stand that was sampled, had an evenly distributed overstory and intense understory regeneration (842 tpa) and competition (mean crown overlap of 72%). It has an overstory SDI of 140, but an average crop tree RVG of only 26% due in large part to the understory SDI of 59. West Wier (Ww, Table 3), in contrast, has an overstory SDI of 223 and a crop tree RVG of 23%, but only has an understory SDI of 7. Understory SDI is a product of both trees per acre and average tree size. Larger understory tree size increases understory SDI and drives down average RVG in the model, which supports the hypothesis that larger trees need more growing space (Givnish 1988, Messier et al. 1999). For example, Rodomsky (Rd) has an overstory SDI of 226, a crop tree tpa of 19, and a crop tree RVG of 16% (Table 3). Somemore (Sm) has a higher overstory SDI of 252, a similar crop tree tpa of 15, but a crop tree RVG of 23%. While low percent shrub cover and a higher SDI plot ratio (ratio overstory SDI of plots with understory trees to SDI of all plots) account for part of this difference, the greater understory tree size in Rodomsky, and thus understory SDI, accounts for 3.5% of the difference. The average tree height is 7.4m, which is high for a stand with such a high level of overstory stocking. Percent cover of tall shrubs substantially reduces understory RVG in the models, indicating the competitive effect that may other authors have shown (Chan and Walstad 1987, Cole and Newton 1987, Wagner and Radosevich 1991, Emmingham et al. 2000, Brandeis et al. 2001b). Low shrubs, on the other hand, increase RVG suggesting that they do not have a net negative competitive effect on trees and are perhaps a sign of greater moisture and/or light reaching the understory. The role of crown overlap and the SDI plot ratio in the model suggests that the horizontal distribution of both understory and overstory has an effect on the relationship of understory vigor and overstory stocking. Other authors have found that for the same amount of overstory stocking, a more clumped overstory distribution with gaps provides more microsites with light levels sufficient for Douglas-fir than a uniform distribution (Bailey and Tappeiner 1998, Lieffers et al. 1999, Coates et al. 2003, Drever and Lertzman 2003). Comparing several stands with similar overstory SDI confirms this (Table 3). Kalipso (KP) has an overstory SDI of 242, an average VOS of 36, and a crop tree RVG of 30% while Cherry Hill (Ch) has an SDI of 163, VOS of 35, and a crop tree RVG of 29%. According to the model, the ability of Kalipso to have such a high RVG for its relatively high amount of overstory stocking is due to low understory stocking (5), low percent cover of tall shrubs (4%) and low SPR (70%). If understory stocking and tall shrubs are increased to levels more similar of other stands with the same level of overstory SDI, crop tree RVG is still 3-8% higher than these other stands. This patchy stand had the majority of its regeneration in gaps while parts of the stand with high overstory densities had no regeneration. The fact that it has such a high average VOS compared to other stands with similar levels of overstory SDI (Table 3) suggest that this clumpy distribution of overstory trees results in a higher average understory light level for similar levels of SDI.
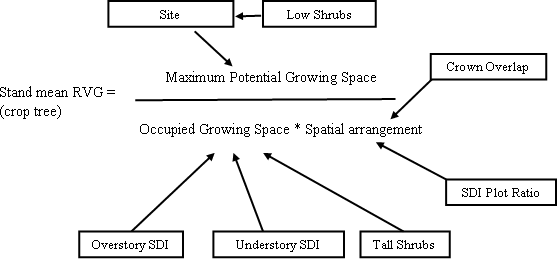
Through sensitivity analysis, the model showed that to achieve an average crop RVG of 35% or a vigor category 3, the highest overstory SDI possible is 210 and the average SDI is 150. For a crop tree RVG of 50%, vigor category 4, the highest SDI is 170, whereas the average is 120. The higher levels of SDI needed to achieve 35 and 50% RVG predicted by the stand model compared to the individual tree model (Figure 7) is due to the fact the stand model is taking into account the overstory in the whole stand and not just overstory trees that are affecting understory trees. All stands had plots with no understory trees that increased average overstory SDI without influencing RVG. Although experimentation with the model did not involve values for predictor variables outside of the ranges found in the stands sampled, actual stands with a crop tree RVG above 36% were not found. The model is not a predictive model, and the results should be used only as general guidelines. The results and overall structure of the model confirm the underpinnings of the stand level, multi-cohort growth model developed by O’Hara (1996) and O'Hara and Valappil (1999). Average vigor of an understory cohort is determined by the growing space available to the cohort divided by the number of trees in the cohort. Using leaf area index to measure growing space, the total available leaf area is apportioned between the overstory and understory cohorts to determine the leaf area available to a particular understory cohort. The model in this study uses SDI to measure growing space occupancy and includes shrubs and the spatial arrangement of the cohorts. The following diagram (Figure 11) explains this concept using the non-linear model form derived in this study.
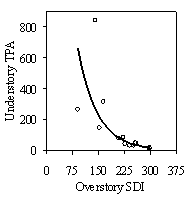
This stand level model was developed from only 13 stands at a single point in time and cannot be used to predict the effects of treatments. The high R2 value of the model is likely to be a product of having only 13 data points and five or six variables. With more stands, the R2 value would likely decrease. However, the fact that between 85-90% of the variation in RVG was predicted with only overstory SDI, understory SDI, and tall or low shrubs, suggests that strong, stand level relationships between understory vigor, overstory and understory SDI, and shrub cover do exist. Until more stands are included, this model should be used as a conceptual model and a rough guide of the quantitative relationships between understory vigor and the predictor variables. Separate from the stand level model, overstory SDI is well correlated with understory stocking, whether measured by tpa or SDI (10b). The exponential relationship suggests that as stands are opened up during future thinnings, natural regeneration will provide sufficient understory stocking levels to maintain adequate growing stock under current thinning regimes. Miller and Emmingham (2001) also found that natural regeneration provided sufficient growing stock in selection thinning systems in similar Douglas-fir forests. However, many gaps in the stands surveyed in this study had low understory stocking and understory crop tree stocking levels are currently on the low side for most of the stands (Table 3). Crop tree stocking should be monitored and a more detailed analysis of growing stock under different thinning regimes should be undertaken to ensure that growing stock is adequate. Conclusions and Management RecommendationsTo assess vigor of understory Douglas-fir, height growth, LC ratio, HD ratio, and crown density rating (a visual estimate or measured variable of crown density, such as number of internodal branches or branch length) can be combined together into a powerful indicator of vigor: relative volume growth (RVG). RVG allows for comparisons across a wide range of size classes, relates easy to measure growth metrics to volume growth, and provides managers with a concrete measure of how understory trees are growing compared to the maximum potential for that site. The qualitative crown density rating can be dropped if necessary with only a small loss in predictive power (Appendix B). Understory tree vigor can also be assessed via a classification system that uses thresholds of height growth, HD ratio, and LC ratio to place trees into four vigor classes. This simpler system is sufficient for most management applications and can be easily integrated into most stand exam, marking, or cruising protocols. With some training, field technicians can quickly classify trees in the field using only visual estimates of these growth metrics, or by taking height and diameter measurements and visually estimating height growth and live crown ratio. It is very likely that most of the Douglas-fir regeneration (vigor classes 1-3) found in this study will need release from overstory removal in order to recruit into the overstory. Although Douglas-fir appears to be able to maintain slow growth rates, reasonable LC ratios and HD ratios, and release potential in low light environments for several decades, it is unlikely that Douglas-fir can persist at slow growth rates in the understory for the many decades it would take for the overstory to be thinned by natural mortality. Understory trees from vigor class one are currently at some risk of mortality from suppression and growth rates of trees from higher vigor classes will decline as the overstory closes over time. If released, however, evidence from this study and all other studies reviewed suggest that Douglas-fir will respond. In all but the most severely suppressed trees, released trees appear able to regain growth rates comparable with trees that were never suppressed, recover from high HD ratios, and become viable replacement trees. The lag time before response to release, the degree of response, and decline in HD ratios after release will depend on the level of overstory removal, the degree of crown crowding from understory competition after release, and pre-release live crown ratio and growth rates. Trees in classes one and two will likely take a period of time to respond, while class three and four will likely respond quickly. Furthermore, risk of stem failure from high HD ratios and lag time before response to release appear to increase with increasing tree height. Trees taller than 5m in both vigor class one and two should be carefully evaluated to determine if they are worth releasing. A LC ratio of at least of 50% and an HD ratio of not more than 80 (measured at 15cm above ground) are suggested thresholds to guide this evaluation. Trees taller than 10m, especially from vigor class 1, are unlikely to be worth releasing.
At the individual tree level, dominant and co-dominant Douglas-fir with low levels of crown crowding (less than 50% 2-dimensional crown overlap) need an average of 45% full sunlight to achieve vigor levels where recruitment into the overstory without further release is likely (vigor class four). For class three trees, this average is 35% full sunlight. For vigor classes one and two, the average was found to be 10-15% full sunlight. Below 10% full sunlight, regeneration is likely to be scarce and in vigor class one. SDI levels are 150 (30% full site occupancy; 24 Curtis Relative Density) or less to achieve vigor classes three and four, 150-275 for classes one and two, and 275+ (55% full site occupancy; 44 Curtis Relative Density) for scarce stocking and class one trees. These percent of full sunlight and SDI levels are means, however, and growth rates of individual trees will vary considerably for a given level of light or overstory density. Trees with high levels of crown crowding (greater than 50% 2-dimensional crown overlap) require more light to achieve the same vigor classes, and this effect increases in higher light environments. Only in very high light environments do they reach 35% RVG (Figure 6 & 7). Even in low light environments, negative effects on HD ratios and live crown ratios are apparent (Figure 9). Without release from crown crowding, trees with greater than 50% crown overlap will suffer serious losses in potential volume growth. Pre-commercial thinning is recommended for crop trees that have crown overlap over 50% or are likely to before the next stand entry. Pre-commercial thinning should be done after overstory thinning treatments as harvesting will damage a proportion of crop trees. While maintaining the overstory below 150 SDI is necessary for recruitment of Douglas-fir advanced regeneration into the overstory, entire stands do not need to be thinned to these low levels. Most volume growth and stand structure exists in the overstory and thus early, heavy thinning across an entire stand will result in a major loss of potential volume growth (O'Hara 1996). It can also result in intense natural regeneration or shrub colonization of open areas. To balance the tradeoff between total stand volume growth and vigor of regeneration, patchy or spatially heterogeneous stands can be created and maintained in a shifting mosaic. A three stage progression of overstory treatment types is recommended for patches within a stand. First, some patches can be managed to maximize volume growth in the overstory for one or more stand entries with light thinning from below treatments that keep stocking levels over 275 SDI. Although some regeneration may establish after these light thinning entries, it should not be a treatment objective as regeneration establishment will be a focus of the second stage. In this second stage, the overstory should be opened up enough (150-275 SDI) for regeneration to establish and maintain at an average vigor of class two. Depending on stand conditions and the objectives of the landowner, overstory trees can be removed across all diameter classes or from below, and small gaps can be created. These two cohort patches can be maintained for several stand entries by progressively reducing overstory density towards 150 SDI which will allow the regeneration to keeping growing, albeit slowly, and maintain its release potential. Depending on how vigorous the regeneration cohort is, the patch should be moved to the third stage once the regeneration is between 5-10m in height. In the third stage, overstory SDI should be reduced below 150 to provide enough light for regeneration to grow vigorously and recruit into the overstory. To maximize growth of the regeneration cohort, all the overstory trees in the patch should be removed in one entry. If a multi-layered canopy is a management goal, however, the overstory can be removed over several stand entries and some overstory trees can be indefinitely retained for long term structure. Clumped retention, rather than dispersed retention, should be used as it provides more micro-sites with high light levels for the same amount of overstory SDI (Bailey and Tappeiner 1998, Lieffers et al. 1999, Coates et al. 2003, Drever and Lertzman 2003) and thus uses growing space more efficiently. A third cohort may establish after scarification from successive stand entries in areas of the patch that are not stocked with regeneration. Once the main regeneration cohort reaches commercial size, the patch can be moved back to the first stage and the cycle can begin again. If some members of the original overstory cohort are retained, a three cohort structure is likely to be achieved in the next cycle if it is not already present.
By ensuring that different patches within a stand are at different stages in the progression, a horizontally and vertically complex stand with multiple cohorts will be created. Although each patch may only have 1-3 cohorts, the patches put together will have more. Minimum patch size should be 0.25 ha (0.6 ac), as this appears to be the minimum gap size necessary for Douglas-fir to achieve high vigor levels in gaps (Ketchum 1994, Mailly and Kimmins 1997). Larger patch sizes, however, will be easier to manage. By varying patch size and the spatial arrangement of patch stages, a high degree of structural complexity can be created. Active management of the regeneration cohort should be also considered during the second and third stages of overstory treatments. As described above, pre-commercial thinning and shrub control around crop trees has the potential to significantly increase vigor levels. While natural regeneration appears to be providing sufficient stocking under thinned understories, gaps captured by shrubs and grasses may need shrub control and planting to ensure timely stocking and full utilization of growing space. Gaps occupied with native shrubs provide important wildlife habitat, but many gaps are filled with non-native Scotch Broom (Cytisus scopariu) in the stands inventoried in this study. Prescriptions for individual stands will need to take into account the current vigor, height, crown crowding, spatial arrangement, and stocking levels of both overstory and understory cohorts, as well as shrub cover. Relatively uniform, even-age stands with high overstory SDI and little regeneration, such as Bad Acre West, will need to be thinned to different densities throughout the stand to create a mix of stage one and two patches. Stage three patches can be added if large gaps are desired for wildlife habitat. Stands that already have a high level of patchiness, such as Kalipso, can be treated by moving different patches to different stages depending on the stocking, vigor and height of the regeneration in each patch. Stage two patches where regeneration is over 5m, such as many areas in Redomsky, should be opened up and moved to stage 3 patches to ensure the recruitment of regeneration into the overstory. Stands, such as Cherry Hill, with overstory stocking close to or below 150 SDI and high levels of understory competition will likely require pre-commercial thinning, in addition to further overstory removal, to move the regeneration into vigor classes three and four. To provide an approximate assessment of stands at the patch level, inventory data can be broken down by plot to show the distribution of overstory SDI and understory trees per acre, average height, relative volume growth, and crown overlap. Displaying this information on stand map using GIS can provide an idea of the spatial distribution of different cohorts (Appendix E). To help managers determine the stand level tradeoffs between structural goals, wood production needs, habitat value, understory vigor, and risk tolerance, stand level models that are based on existing inventory information and accessible to managers are needed (Lieffers et al. 1999, Hasenauer and Kindermann 2002). While light based models are clearly superior at predicting growth of advanced regeneration at the individual tree level, density based models appear to work well at the stand level (O'Hara 1996). Although spatially explicit models are powerful tools to understand forest ecosystems and the effects of experimental treatments, they will require substantial investment of time and resources to develop and collect spatially explicit inventory information on a broad scale is not realistic at this point in time. At the same time, traditional inventory metrics and growth models designed for even-age management are often not sufficient for uneven-age management. Incorporating light measurements and simple metrics for intra-cohort competition, spatial distribution of cohorts, and shrub cover into distant independent stand models shows promise to make them versatile and powerful enough for most management needs. The results of this study support the conclusions of other investigators that uneven-age management is possible with Douglas-fir on dry sites using two-aged shelterwood systems (Seidel 1983b, Tesch and Korpela 1993), group selection (Coates and Burton 1997), or single tree selection systems that include heavy thinning at some point (Deisenhofer 2000, Miller and Emmingham 2001). By combining elements of all three systems, structurally complex, multi-cohort stands can be created that also produce significant wood volume. These stands will be structurally different from west-side, late-successional forests that contain shade tolerant conifers, however, and require ongoing, periodic thinning to maintain.
APPENDICESAppendix A
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||